Skip to content
buzardweb@gmail.com
Pinterest-p
500px
Flickr
Rss
Home
About
Categories
Animals, Aquariums and Zoos
Arabia
Archaeology
Arizona
Australia
Belgium
Birds
Botswana
Bridges and Buildings
British Museum
Bugs, Bees and Butterflies
California
Canada and Alaska
Cathedrals & Churches
Clocks and Astrolabes
Colombia
Colorado
Cooking and Recipes
Costa Rica
Decorative Arts
Egypt
Ethiopia
Events
Fashion and Shopping
Food and Drinks
France
Paris
History
Hotels and Lodges
Israel
Italy
Jewels and Jewelry
Jordan
Las Vegas
Loire Valley
London
Madagascar
Maps
Monument
Museums
Myth, Philosophy and Religion
Native Americas Cultures
New Zealand
Painting
Peru
Photographic Equipment and Techniques
Plants and Gardens
Portugal
Reims
Restaurant Reviews
Rome
Rouen and Normandy
Sculpture
South Africa
The Louvre
This and That
Travel Tips
Trinidad and Tobago
Turkey
Versailles
Vienna
Washington DC
Weapons and War
Blog
Galleries
Menu
Home
About
Categories
Animals, Aquariums and Zoos
Arabia
Archaeology
Arizona
Australia
Belgium
Birds
Botswana
Bridges and Buildings
British Museum
Bugs, Bees and Butterflies
California
Canada and Alaska
Cathedrals & Churches
Clocks and Astrolabes
Colombia
Colorado
Cooking and Recipes
Costa Rica
Decorative Arts
Egypt
Ethiopia
Events
Fashion and Shopping
Food and Drinks
France
Paris
History
Hotels and Lodges
Israel
Italy
Jewels and Jewelry
Jordan
Las Vegas
Loire Valley
London
Madagascar
Maps
Monument
Museums
Myth, Philosophy and Religion
Native Americas Cultures
New Zealand
Painting
Peru
Photographic Equipment and Techniques
Plants and Gardens
Portugal
Reims
Restaurant Reviews
Rome
Rouen and Normandy
Sculpture
South Africa
The Louvre
This and That
Travel Tips
Trinidad and Tobago
Turkey
Versailles
Vienna
Washington DC
Weapons and War
Blog
Galleries
Search
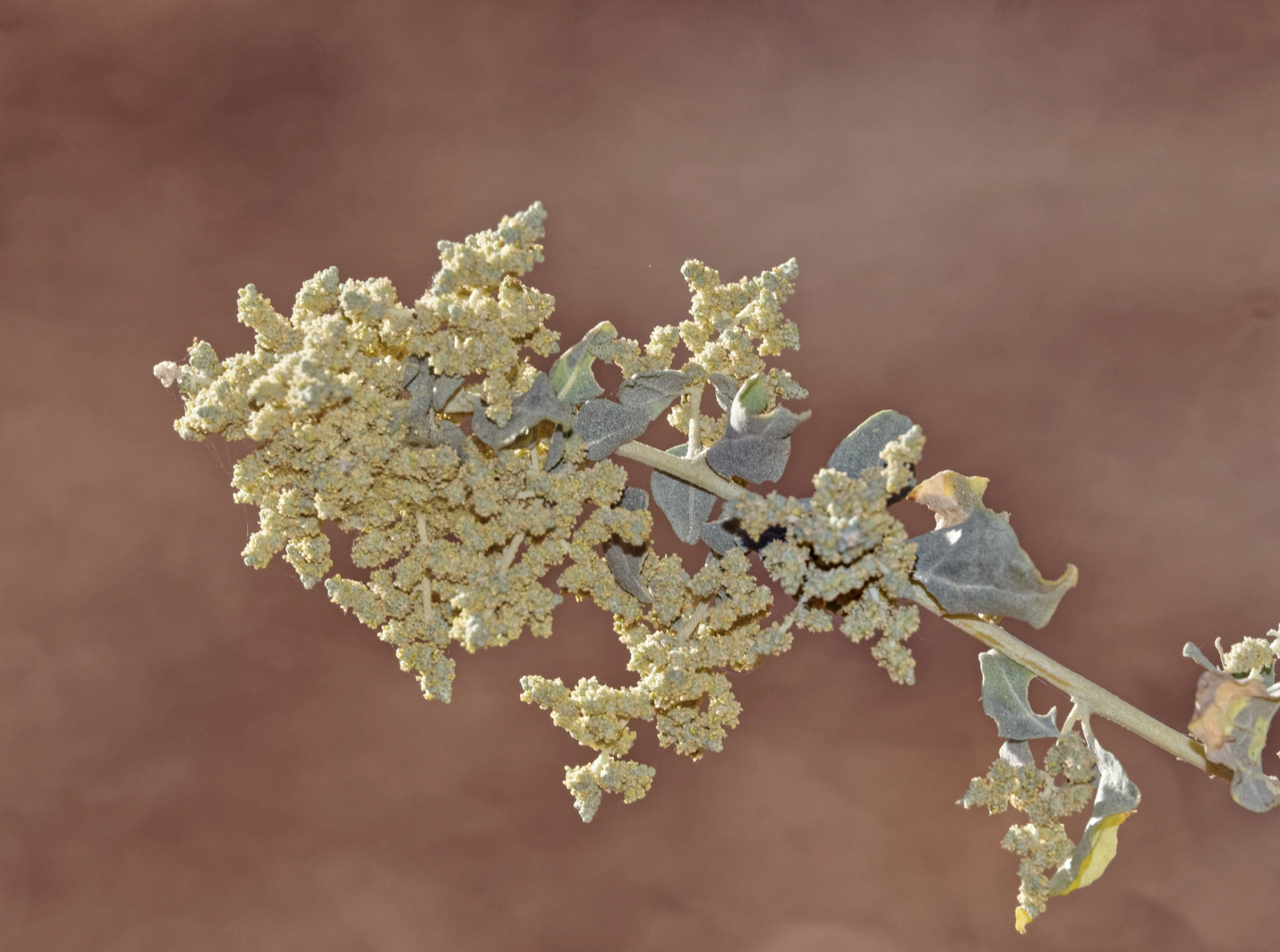
Desert Adaptations
Home
»
Desert Adaptations
Stem Photosynthesis in Select Mohave Desert Plants
May 25, 2021
Desert Adaptations in Mohave C4 Plants
March 6, 2021