
This post is the second on my series on Mohave desert plants, this time focused on C4 plants, not really as complex as it might sound. The C4 photosynthetic pathway has evolved an estimated 45 times in terrestrial plants (Sage 2004), and is most prominent in grasses, which account for roughly 25% of global terrestrial primary production (Still et al. 2003) and include important crop and weed plants and potential biofuels such as maize, sugarcane, sorghum and switchgrass. The highest rate of photosynthesis is typically observed in C4 plants. The photosynthetic rate in such plants is known to be directly related with the variation of the solar rays in the daytime. Maximum rate of photosynthesis occurs in the red and blue regions of the visible light as seen in the absorption spectra of chlorophyll a and b. These are of economic importance as they have a comparatively higher photosynthetic efficiencies in comparison to other plants. C4 photosynthetic plants outperform C3 plants in hot and arid climates. By concentrating carbon dioxide around Rubisco C4 plants drastically reduce photorespiration. The frequency with which plants evolved C4 photosynthesis independently challenges researchers to unravel the genetic mechanisms underlying this convergent evolutionary switch. The conversion of C3 crops, such as rice, towards C4 photosynthesis is a long‐standing goal. Nevertheless, at the present time, in the age of synthetic biology, this still remains a monumental task, partially because the C4 carbon‐concentrating biochemical cycle spans two cell types and thus requires specialized anatomy.
Photorespiration and the Enzyme RuBisCO
The majority of plant species on Earth uses C3 photosynthesis, in which the first carbon compound produced contains three carbon atoms. In this process, carbon dioxide enters a plant through its stomata (microscopic pores on plant leaves), where amidst a series of complex reactions, the enzyme RuBisCO fixes carbon into sugar through the Calvin-Benson cycle. However, two key restrictions slow down photosynthesis.
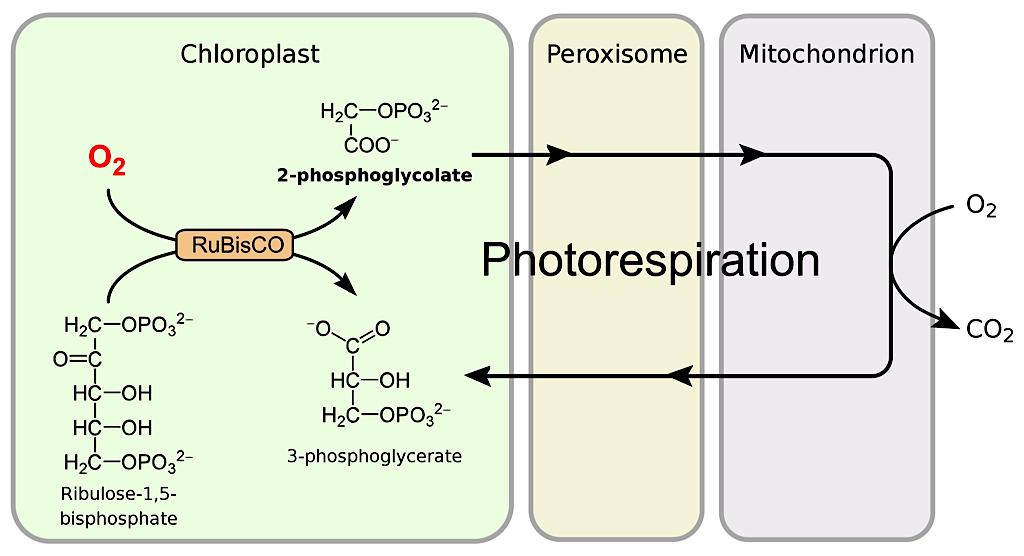
RuBisCO is intended to fix carbon dioxide, but it can also fix oxygen molecules, which creates a toxic two-carbon compound (C2 carbon fixation). RuBisCO fixes oxygen about 20 percent of the time, initiating a process called photorespiration that recycles the toxic compound. Photorespiration costs the plant energy it could have used to photosynthesize. When stomata are open to let carbon dioxide in, they also let water vapor out, leaving C3 plants at a disadvantage in drought and high-temperature environments. However, plants have evolved another form of photosynthesis to help reduce these losses in hot, dry environments.
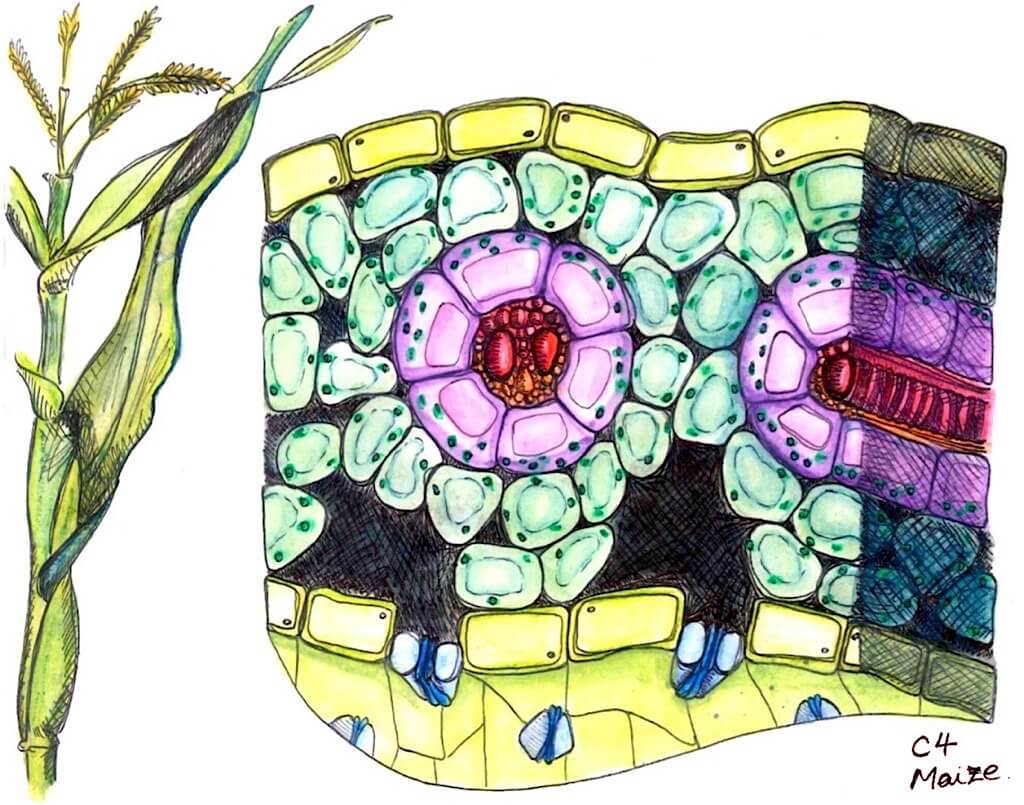
In C4 photosynthesis, where a four-carbon compound is produced, unique leaf anatomy allows carbon dioxide to concentrate in ‘bundle sheath’ cells around RuBisCO. This structure delivers carbon dioxide straight to RuBisCO, effectively removing its contact with oxygen and the need for photorespiration. What’s more, this adaptation allows plants to retain water through the ability to continue fixing carbon while stomata are closed. Although most C4 plants exhibit Kranz anatomy, there are, however, a few species that operate a limited C4 cycle without any distinct bundle sheath tissue. C4 plants are often identified by microscopic slide cross sections showing this distinctive Kranz anatomy.
Carbon Isotope Ratio and Photosynthetic Pathways
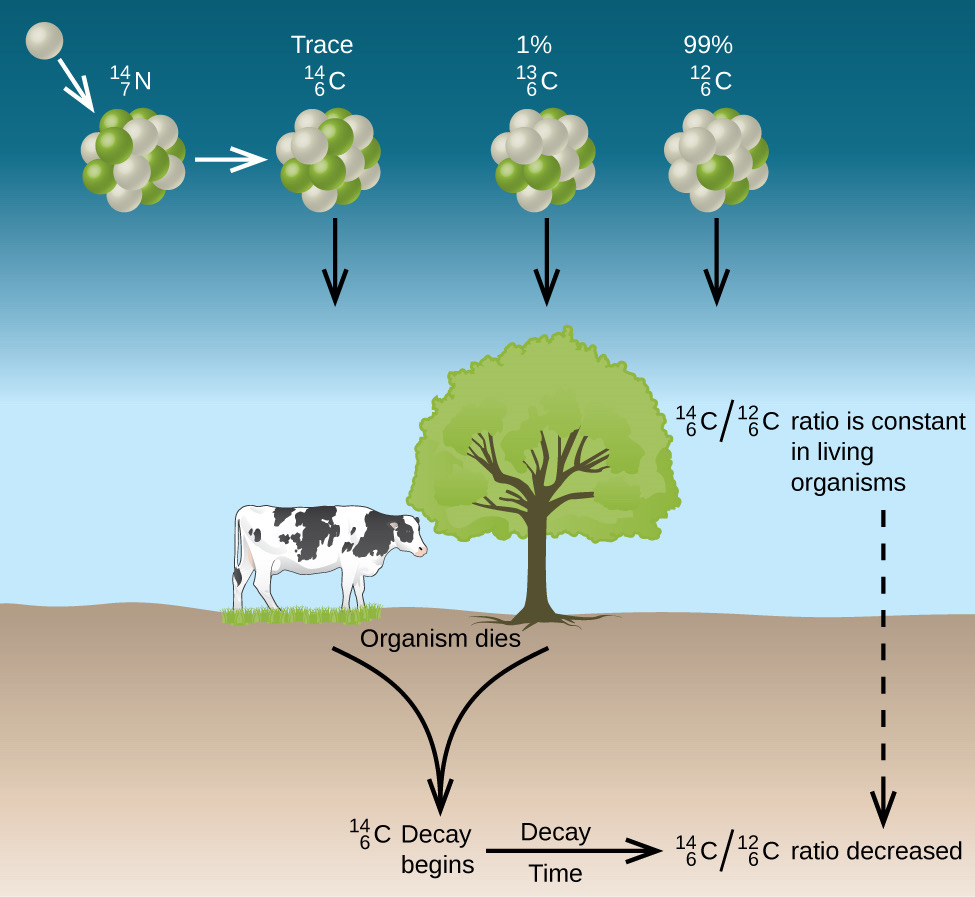
There are three naturally occurring isotopes of carbon: 12, 13, and 14. 12C and 13C are stable, occurring in a natural proportion of approximately 93:1. 14C is produced by thermal neutrons from cosmic radiation in the upper atmosphere, and is transported down to earth (as CO2) to be absorbed by living biological material. Isotopically, 14C constitutes a negligible part; but, since it is radioactive with a half-life of 5,700 years, it is radiometrically detectable. Since dead tissue does not absorb 14C, the amount of 14C is one of the methods used within the field of archeology for radiometric dating of biological material.
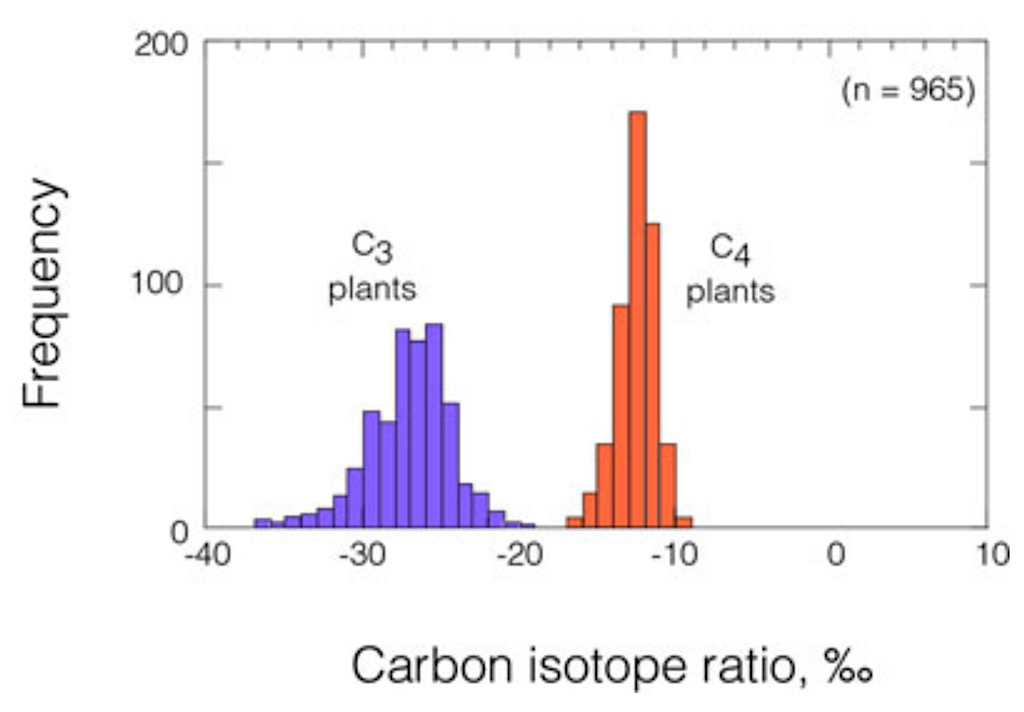
Variations in the carbon isotope ratios of C3 and C4 plants. Cerling et al. 1997The amount of 12C and 13C in plant tissue can be measured by mass spectrometry and compared to a standard (the ratio in the air); the result (e.g. the delta of the 13C = δ13C) is expressed as parts per thousand. This comparison between the relative amounts of 12C and 13C is usually computed as a ratio called the “Carbon Isotope Ratio”.
Since the initial observations of Bender (1968, 1971) and Smith and Epstein (1971), there has been an interest in using carbon isotope ratios as an means of screening plants for C3 versus C4 photosynthetic pathway differences. It also became clear that plants with Crassulacean acid metabolism (CAM) exhibited carbon isotope ratios similar to C4 plants or in some species had carbon isotope ratios intermediate between those of C3 and C4 plants (Osmond ct al. l982). We now know that the carbon isotope ratios of leaves can vary from –7 to –35%, with C4 plants having values of –7 to –15%, CAM plants –10 to –22%, and C3 plants –20 to –35%. The source for this discrimination in C3 photosynthesis is the initial carboxylating enzyme ribulose hisphosphate (RuBP) carboxylase, which discriminates strongly against the heavier isotope, 13C. Phosphoenolpyruvate (PEP) carboxylase (as in C4 and CAM species) appears not to discriminate against 13C, but diffusional and recarboxylation factors play an important role in determining the isotopic composition. Little further discrimination occurs after photosynthesis within the leaf, except during lipid metabolism (DeNiro and Epstein I977). Carbon isotope ratio surveys to determine the presence and abundance of C3, C4 and CAM photosynthetic pathways have been made throughout the world in arid deserts. Several general patterns have emerged from these surveys. First. although C3, C4 and CAM photosynthetic pathways occur in plants from each of these aridland regions. it appears that the greatest fraction of the non-succulent species are C3 plants. The succulent species tend to be obligate-CAM or facultative-CAM plants, with the distinction correlated with stem versus leaf succulence. When the C4 photosynthetic pathway occurs. it is most frequent among perennial halophytes (salt-tolerant plants like Atriplex) and annuals. There is also a correlation between photosynthetic pathway and precipitation, so that C3 plants tend to predominate at the mesic (well watered) sites while CAM plants are most frequent at the driest locations. In nonsaline regions, C4 plants tend to predominate only on those arid locations where there is significant summer precipitation or a source of water such as a stream, seep or spring.
Shadescale (C4)



Atriplex confertifolia, the shadscale, is a species of evergreen shrub in the family Amaranthaceae, which is native to the western United States and northern Mexico. Shadscale fruits and leaves provide important winter browse for domestic livestock and native herbivores. Compared to fourwing saltbush (Atriplex canescens), shadscale has shorter and wider leaves and the fruit does not have four wings (although it may have two wings in a “V” shape). As with other desert climate members of the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells.As with other desert climate members of the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells. Many desert plants have hairy leaves or stems. The hairs act like a blanket over the leaves to protect them from the heat and cold. The white color reflects the sunlight helping to keep the plant cool. They also help trap any moisture and reduce the amount of water lost.
Desert Holly (C4)


Desert Holly (Atriplex hymenelytra). Rogers Spring Near Overton, NevadaDesert holly grows in alkaline locations such as desert dry wash and creosote bush scrub in the Mojave Desert and Sonoran Desert down to Baja California. It is generally a compact, rounded bush, 8–48 inches (20–122 cm) tall, covered in distinctive reflective silver-gray, twisted, oblong, many-pointed leaves. It drops its leaves drought deciduous in extreme drought conditions. It tolerates alkaline soil, salt and sand. The leaves accumulate salts which helps extract water from the soil when other plants cannot. Salt is shed by dropping the leaves. As with other desert climate members of the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells.
Fourwing Saltbush (C4)


Fourwing saltbush is a nondescript shrubby member of the goosefoot family (Chenopodiaceae) that ranges across much of the southern Great Plains, Great Basin, and desert southwest. It is characterized by grayish, linear to spoon-shaped leaf blades, yellowish twigs, and small, yellowish, or brown wind-pollinated flowers clustered in slender inflorescences and lacking showy petals. Mature fruits can be eye-catching because of their relatively large size (1/2 to 1 inches long and broad) and four membranous wings (thus, the name “fourwing”). Fourwing saltbush is often abundant, especially in sandy areas, and easy to ignore. The Native American Hopi Indians preferred the ashes of four-wing saltbush for the nixtamalization of maize (the first step in the process of creating tortillas and pinole, by which the pericarp of Indian corn is removed before parching and grinding). The process of nixtamalization was first developed in Mesoamerica, where maize was originally cultivated. The Pima Indians used to eat the seeds. Native Americans of the Southwest cooked the seeds of the four-wing saltbush like oatmeal, and they would eat the leaves either raw or cooked. The plant is rich in niacin, so it was very important to their diet. Navajos made the twigs and leaves into a yellow dye. As with other desert climate members of the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells.
Quail Bush (C4)



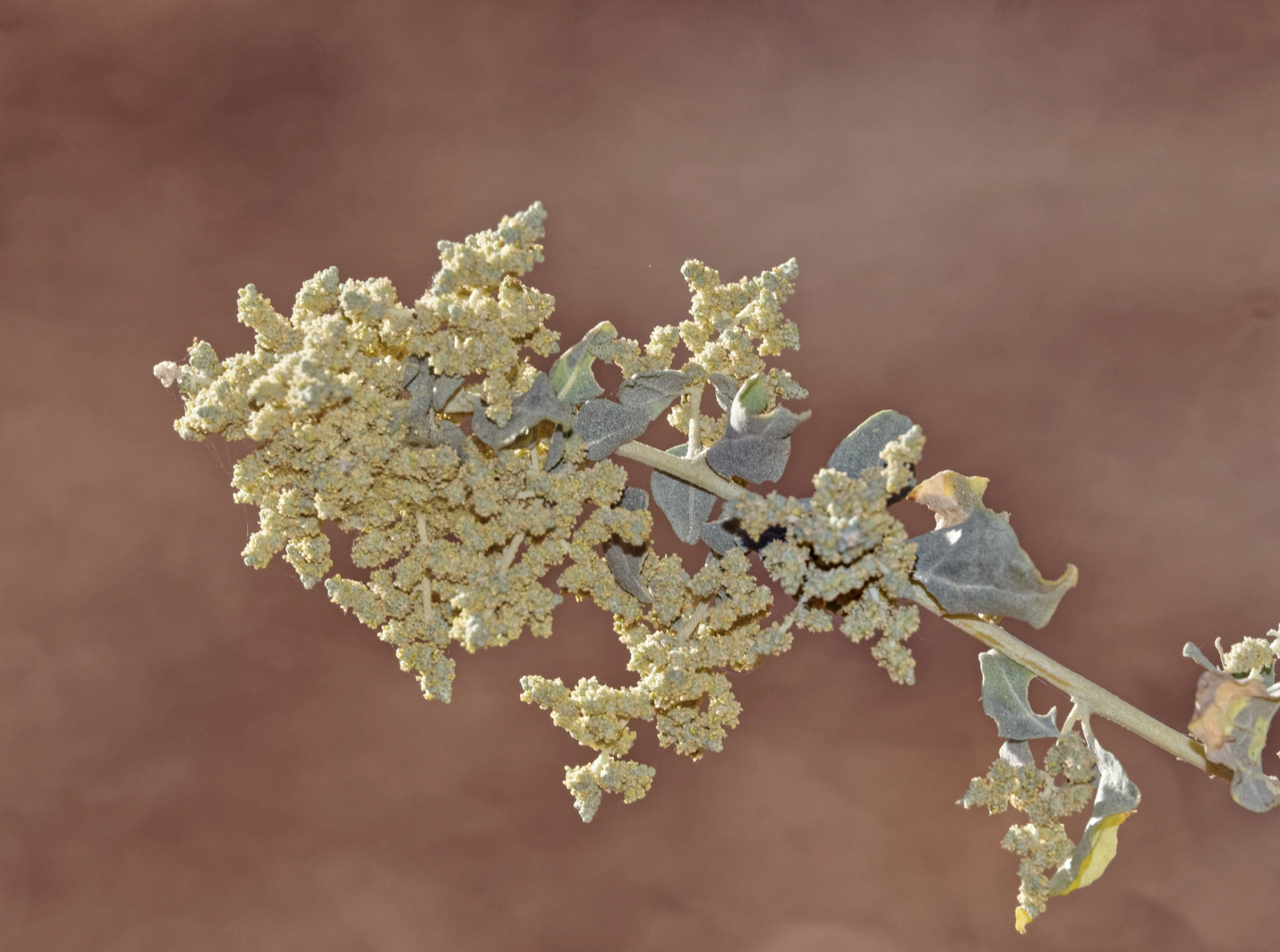

Atriplex lentiformis (quail bush, big saltbrush, big saltbush, quailbrush, lenscale, len-scale saltbush and white thistle) is native to the southwestern United States and northern Mexico, where it grows in habitats with saline or alkaline soils, such as salt flats and dry lake beds, coastline, and desert scrub. It can also be found in nonsaline soils on riverbanks and woodland. Despite the ability of big saltbush to tolerate drought, establishment and survival are probably improved with greater availability of water. Young big saltbush plants have been reported to be more susceptible to drought. Atriplex lentiformis is a spreading, communal shrub typically reaching 3–6 feet (1–2 m) in height and generally more in width. It is highly branched and bears scaly or scurfy gray-green leaves up to 2 inches (5 cm) long and often toothed or rippled along the edges. This species may be dioecious or monoecious, with individuals bearing either male or female flowers, or sometimes both. Male flowers are borne in narrow inflorescences up to 20 inches (50 cm) long, while inflorescences of female flowers are smaller and more compact. Plants can change from monoecious to dioecious and from male to female and vice versa. The mature seeds are red. As with other desert climate members of the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells.
Salt Heliotrope (C4)




Heliotropium curassavicum, commonly called salt heliotrope, a species of flowering plant in the borage family (Boraginaceae). It is native to much of the Americas, from Canada to Argentina, including the West Indies and Hawaii. Due to its wide geographical range that spans many nations and languages, Heliotropium curassavicum has been given an assortment of common names. In English, these include seaside heliotrope, salt heliotrope, monkey tail, quail plant and Chinese parsley. In Latin American Spanish, it is known as cola de mico, cola de gama or rabo alacrán. It is called kīpūkai in Hawaii. It can be found as an introduced, and sometimes invasive, species in Africa, Asia, Australia, and Europe. It thrives in salty soils, such as beach sand, alkali flats, and salt marshes. It is often found in disturbed coastal sites. This is a perennial herb which can take the form of a prostrate creeper along the ground to a somewhat erect shrub approaching 1.5 feet (0.5 m) in height. The stem and foliage are fleshy, with the leaves thick and oval or spade-shaped. The plentiful inflorescences are curled, coiling double rows of small bell-shaped flowers. Each flower is white with five rounded lobes and a purple or yellow throat. The fruit is a smooth nutlet. There are five currently recognized varieties, Heliotropium curassavicum var. obovatum is the one most often found in Nevada. As with the genus Atriplex, it uses water conserving C4 photosynthesis, and it removes salts by having bladders in the leaves that keep the salt from the plant cells.
Bush Seepweed (C4)


Bush Seepweed or Mojave Sea-Blite Seeds (Suaeda nigra). WikipediaSuaeda nigra, often still known by the former name Suaeda moquinii, is a species of flowering plant in the amaranth family, known by the vernacular names bush seepweed or Mojave sea-blite. It is native to much of western North America, from central Canada through the Western United States, Great Basin, the Mojave Desert in California, and into northern Mexico. The species is a shrub or subshrub growing from a woody base with many spreading branches, reaching up to 1.5 metres (4.9 ft) in height. The plants may be facultative annuals when they find themselves growing in seasonally flooded wetlands. It may be hairy to hairless but is usually waxy. It may be green to red to dark purple in color, sometimes almost black. The succulent leaves are linear to lance-shaped, 1 to 3 centimetres long, and flat or cylindrical. Flowers occur in clusters along the upper stems, each cluster containing 1 to 12 flowers. Leaf-like bracts accompany the clusters. The flower has no petals and is composed of a calyx of fleshy, rounded sepals. As with the genus Atriplex, it uses water conserving C4 photosynthesis.
Desert Saltgrass (C4)


Distichlis spicata is a species of grass known by several common names, including seashore saltgrass, inland saltgrass, and desert saltgrass. It thrives along coastlines and on salt flats and disturbed soils, as well as forest, woodland, montane, and desert scrub habitats. This grass is native to the Americas, where it is widespread. It can be found on other continents as well, where it is naturalized. It is extremely salt tolerant. Native Americans that inhabited California used saltgrass as a seasoning. They collected the salt crystals by threshing the blades. The seasoning provided is gray-green and said to have tasted like a salty dill pickle. This grass is a good example of the difficulties of defining the photosynthetic pathway of a particular species. There are literally thousands of species of C3, C4 and intermediate species of grass, let alone other types of plants. In this case I found a paper by Cloern et al that identifies the carbon isotope ratio of Distichlis spicata as –15, well within the range of C4 plants.
C4 Pathway in Tropical Grasses
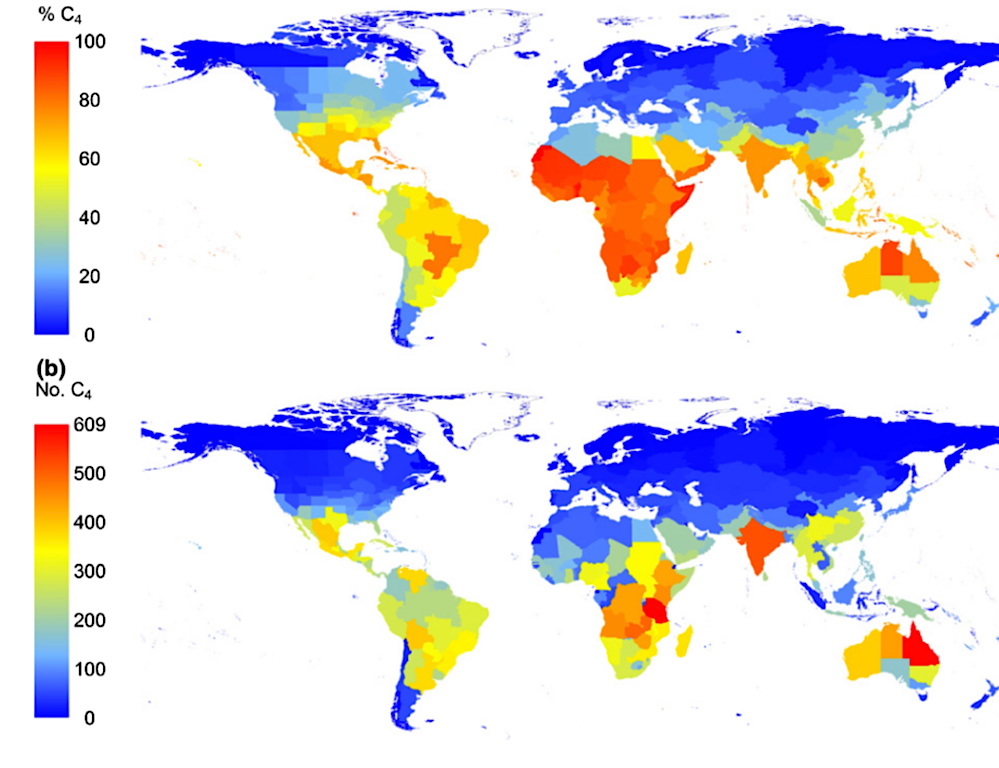
As I wrap up this post, I leave you with this map of C4 grass species, clearly showing that C4 photosynthesis in grasses is more common in hot climates. The paper also gives a list of verified C4 pathway grasses.
As always I hope you have enjoyed the post and will return for more in the future. I realize that the subject matter is dense but I encourage you to enjoy the photos if nothing else. These posts are the framework for future posts on improving food security in a world with too many people.
References:
Desert Adaptations in Mohave C3 Plants
Photorespiration and the Evolution of C 4 Photosynthesis
Annual Plants: Adaptations to Desert Environments
Carbon Reduction Pathways and Standing Crop in Three Chihuahuan Desert Plant Communities
Roles of the bundle sheath cells in leaves of C3 plants
Carbon Isotope Ratios in C3, C4 and CAM Plants
Distichlis spicata Carbon Isotope Ratio –15. Cloern et al
SALT MARSH GRASS, DISTICHLIS SPICATA