

This is the final post in my series on photosynthesis in Mohave desert plants. While green leaves are commonly considered as the primary sources of photosynthesis, higher plants can potentially use almost all vegetative and reproductive structures to perform carbon fixation. Chlorophyll-containing bark and wood tissue, most fruit, root and fertile flower organs typically perform an effective internal CO2 recycling using the respiratory released CO2. Photosynthetic stems have positive effects on the carbon economy of plants through two main mechanisms. Photosynthetic stems can either assimilate atmospheric CO2 and contribute to the net carbon gain of the plant through the process of stem net photosynthesis, or decrease respiratory losses by recycling CO2 previously respired by roots and stems through the process of stem recycling photosynthesis.Plants with green stems are categorized by one of three types of stem photosynthesis syndromes. Retamoids include leafless or almost leafless woody plants that have stomata in the stem epidermis allowing for gas exchange with the atmosphere (Schaedle 1975). Another type of photosynthetic stems are the succulent pachycauls or the sarcocaulescent group, which has large-sized stems with translucent exfoliating bark, a large amount of parenchymatous tissue that serves as a water reservoir, and non-succulent, drought-deciduous leaves (Franco-Vizcaino et al.1990). An example of this would be the Boojum Tree (Fouquieria columnaris) and the related Ocotillo (Fouquieria splendens) as both lack stomata and recycle respiratory released CO2 to survive during drought. Another type of photosynthetic stem is found in cactus, which take up carbon dioxide at night with stoma using CAM photosynthesis. The inadequacy of the current state of knowledge for describing or understanding the diversity of structure, function, and ecological significance of photosynthetic stems suggest areas for further research. The diversity of taxonomic types, and classes of photosynthetic stems, results in a large diversity of structural characteristics. In this post, I have simply chosen interesting examples without resolving these difficulties. I have also decided to throw caution to the wind and use lots of technical jargon.
Xylem and Phloem
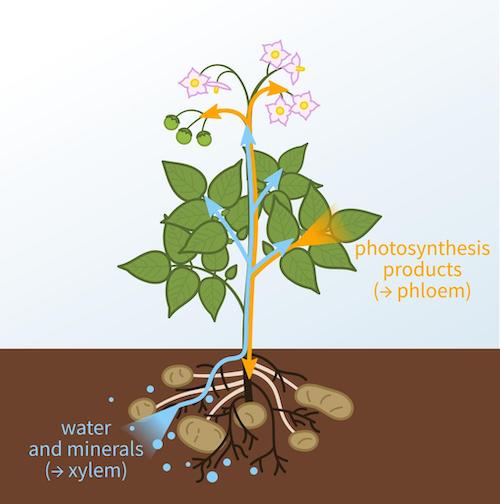
The xylem is responsible for keeping a plant hydrated. Xylem sap travels upwards and has to overcome serious gravitational forces to deliver water to a plant’s upper extremities, especially in tall trees. This is accomplished by a combination of root pressure (pushing up) and transpiration at the leaves (pulling up). Xylem cells are dead, elongated and hollow. They have secondary cell walls and ‘pits’ (areas where the secondary cell wall is missing). Clem forms vascular bundles with phloem and gives mechanical strength to plant due to presence of lignin cells. The lignified secondary wall also makes the xylem waterproof and prevents it from collapsing under the pressure of water transpiration. Phloem transports food and nutrients such as sugar and amino acids from leaves to storage organs and growing parts of plant. This movement of substances is called translocation. In trees, the phloem is the innermost layer of the bark. Phloem tissue consists of conducting cells, generally called sieve elements, parenchyma cells, including both specialized companion cells or albuminous cells and unspecialized cells and supportive cells, such as fibers and sclereids.
Vascular Bundles
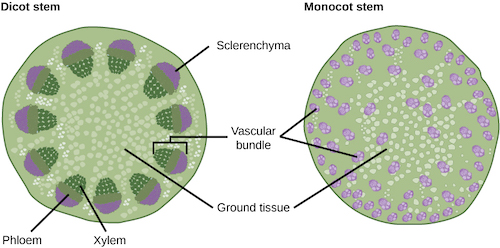
The monocots or monocotyledons have, as the name implies, a single (mono-) cotyledon, or embryonic leaf, in their seeds. Historically, this feature was used to contrast the monocots with the dicotyledons or dicots which typically have two cotyledons; however modern research has shown that the dicots are not a natural group, and the term can only be used to indicate all angiosperms that are not monocots and is used in that respect here. From a diagnostic point of view the number of cotyledons is neither a particularly useful characteristic (as they are only present for a very short period in a plant’s life), nor is it completely reliable. The single cotyledon is only one of a number of modifications of the body plan of the ancestral monocotyledons, whose adaptive advantages are poorly understood, but may have been related to adaption to aquatic habitats, prior to radiation to terrestrial habitats.
Woody Plants
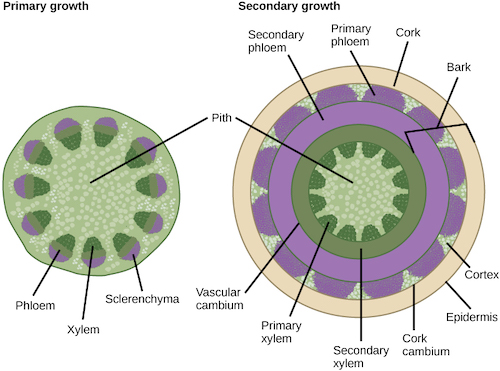
In woody plants, cork cambium is the outermost lateral meristem. It produces cork cells (bark) containing a waxy substance known as suberin that can repel water. The bark protects the plant against physical damage and helps reduce water loss. The cork cambium also produces a layer of cells known as phelloderm, which grows inward from the cambium. The cork cambium, cork cells, and phelloderm are collectively termed the periderm. The periderm substitutes for the epidermis in mature plants. In some plants, the periderm has many openings, known as lenticels, which allow the interior cells to exchange gases with the outside atmosphere. This supplies oxygen to the living and metabolically active cells of the cortex, xylem and phloem.
Stomata vs Lenticels

Lenticels are lens-shaped spots or present in the woody trunks or stems of the plants. They act as pores which involve mainly in the direct gas exchange of plants between internal cells of the stem and the environment. Lenticels appear as spongy areas on plant stems. The shape of the lenticels differs according to the type of the plant. Hence, the shape of the lenticel is one of the characteristics of the plant and can be used as a parameter for tree identification. During the night time, gas exchange occurs through lenticels. Lenticels are always open. They cannot be closed whenever needed. And also they are unable to photosynthesize due to the absence of chlorophylls. Lenticels are visible on surfaces of fruits such as mango, apple and avocado. Lenticels permit exchange of gases between the environment and the internal tissue spaces of the organs. They permit the entrance of oxygen and simultaneously the output of carbon dioxide and water vapor. In most fruits, lenticels account for up to 23% of transpiration.
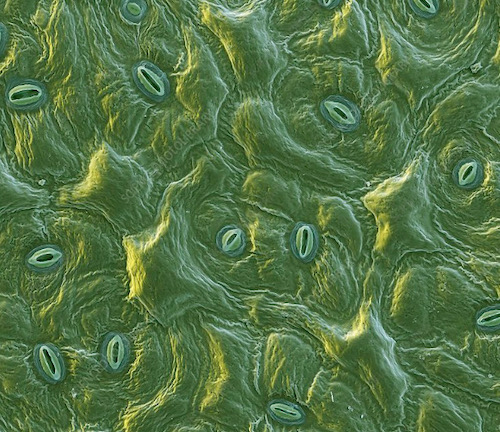
Stomata are normally found in plant leaves but can also be found in some stems. Specialized cells referred to as guard cells surround stomata and function to open and close stomatal pores. Stomata are spores found in the epidermis of leaves, stem and other organs that are used to control gas exchange. They guard cells help plants to reduce water lose to the environment when conditions are hot and dry by ensuring that the stomata remains closed. More importantly, stomata allow plants to take in carbon dioxide, which is necessary for photosynthesis. Terrestrial plants typically have thousand of stomata on the surfaces of their leaves. Majority of stomata are located on the underside of plant leaves reducing their exposure to heat and air current. In desert and aquatic plants, stomata are located on the upper surface of the leaves.
Corticular Stem Photosynthesis
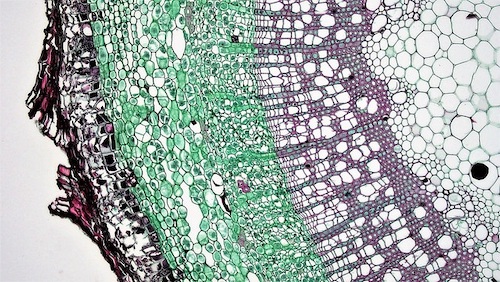
Below the outer peridermal or rhytidomal layers, most stems of woody plants possess greenish tissues. These chlorophyll-containing tissues (the chlorenchymes) within the stems are able to use the stem internal CO2 and the light penetrating the rhytidome to photoassimilate and produce sugars and starch. Although net photosynthetic uptake of CO2 is rarely found, stem internal re-fixation of CO2 in young twigs and branches may compensate for 60–90% of the potential respiratory carbon loss. Isolated chlorenchymal tissues reveal rather high rates of net photosynthesis (being up to 75% of the respective rates for leaf photosynthesis). Corticular photosynthesis is thus thought to be an effective mechanism for recapturing respiratory carbon dioxide before it diffuses out of the stem. Furthermore, chloroplasts of the proper wood or pith fraction also take part in stem internal photosynthesis.
Tree Tobacco (C3 leaves and C4 Retamoid Stem)



Nicotiana glauca is a species of wild tobacco known by the common name tree tobacco. Its leaves are attached to the stalk by petioles (many other Nicotiana species have sessile leaves), and its leaves and stems are neither pubescent nor sticky like Nicotiana tabacum. It resembles Cestrum parqui but differs in the form of leaves and fusion of the outer floral parts. It grows to heights of more than 6 feet. The most common tobacco in the west and southwest is Nicotiana glauca, commonly called tree tobacco or Indian tobacco. It’s a member of the Nightshade family and not a native of the U.S., but was introduced first into California by the Spanish missionaries from Bolivia, Argentina, and Paraguay. The plant is used for a variety of medicinal purposes and smoked by Native American groups. The Cahuilla people used leaves interchangeably with other tobacco species in hunting rituals and as a poultice to treat swellings, bruises, cuts, wounds, boils, sores, inflamed throat, and swollen glands. It is poisonous to eat any part of this plant. Although tree tobacco does contain about one percent nicotine, it also has about 10 percent of anabasine, the alkaloid found in its leaves, stems and flowers. This alkaloid has been linked to birth defects in cows, pigs, and sheep who graze on the plant.
C4 Stem Photosynthesis in Tobacco Plants
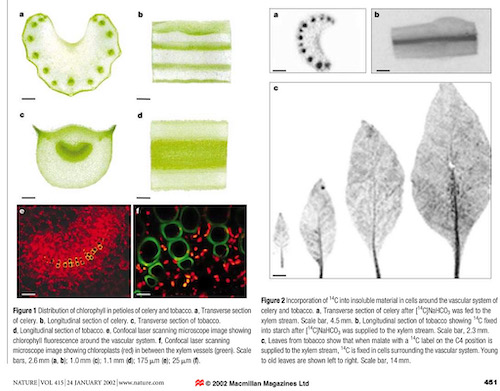
One of the best known and most studied examples of C4 stem photosynthesis is found in the stem of tobacco plants. In fact, all green stemmed plants and virtually all other plants perform photosynthesis in their stems, clustered around the vascular cells. Most of the CO2 comes from internal photorespiration, but may also enter via stem stomata or lenticules. Consider tobacco plants as a template of other plants.
“Most plants are known as C3 plants because the first product of photosynthetic CO2 fixation is a three-carbon compound. C4 plants, which use an alternative pathway in which the first product is a four-carbon compound, have evolved independently many times and are found in at least 18 families. In addition to differences in their biochemistry, photosynthetic organs of C4 plants show alterations in their anatomy and ultrastructure. Little is known about whether the biochemical or anatomical characteristics of C4 photosynthesis evolved first. Tobacco, a typical C3 plant, shows characteristics of C4 photosynthesis in cells of stems and petioles that surround the xylem and phloem, and that these cells are supplied with carbon for photosynthesis from the vascular system and not from stomata. These photosynthetic cells possess high activities of enzymes characteristic of C4 photosynthesis, which allow the decarboxylation of four-carbon organic acids from the xylem and phloem, thus releasing CO2 for photosynthesis. These biochemical characteristics of C4 photosynthesis in cells around the vascular bundles of stems of C3 plants might explain why C4 photosynthesis has evolved independently many times.”
Tobacco C3 Leaves and C4 Stems. Julian M. Hibberd & W. Paul Quick
Palo Verde Tree (C3 Leaves and C4 Retamoid Stem)




Parkinsonia florida (syn. Cercidium floridum), the Blue Palo Verde is primarily found in the Sonoran Colorado Desert of southeastern California, and the Sonoran Deserts of southern Arizona and of northwestern Sonora state (Mexico). Also found in the far eastern Mojave Desert of California in the northern Lower Colorado River Valley, and occasionally in the Mojave’s mountains. Its name means “green pole or stick” in Spanish, referring to the green trunk and branches, that perform photosynthesis. It is found predominantly in desert washes or bajadas, a result of its need for water, although occasionally it can be found in creosote desert scrub habitat, accessing seeps in desert hills up to 3,600 feet (1,100 m).


These trees can photosynthesize through their green bark, an important adaptation for a tree that drops its leaves during the warm season and in response to fall cooling. Palo verdes also drop stems and branches to combat drought. Blue Palo Verde has a deep root penetration, microphyllous leaves, hair cell adornment, decumbent trunks, drought-deciduous behavior, petiole, stem, and trunk photosynthesis, thick cuticular covering on petioles, stems and trunks, sunken stoma, C4 photosynthesis, deep penetration of solar radiation into lignified stems, and trunks, (and vascular systems, which transmit the light deep into trunk tissues), lignified xylem to prevent cavitation, and production and storage of oils and starch as well as druse crystals of calcium oxalate. Palo verde is generally found as a small tree or shrub, which lives across the dry Sonoran and Arizona deserts. The tree has a very deep root system, which allows it to locate and use deep groundwater and to survive extended periods of drought. The generous root system also allows it to survive intermittent flash floods, which are common in the southwestern deserts during summer months. Trunks may approach 10in (25cm) in diameter and may be decumbent or vertical. Most palo verde are shrubs, approximately 3 feet tall and 3 feet in width.
Palo Verde (C3 with C4 Retamoid Stem) Microscopy
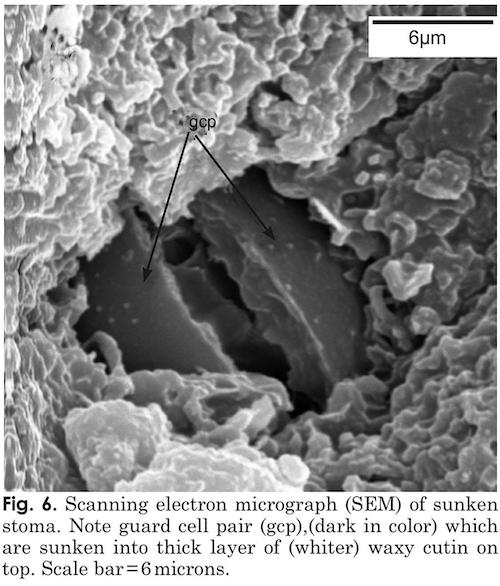
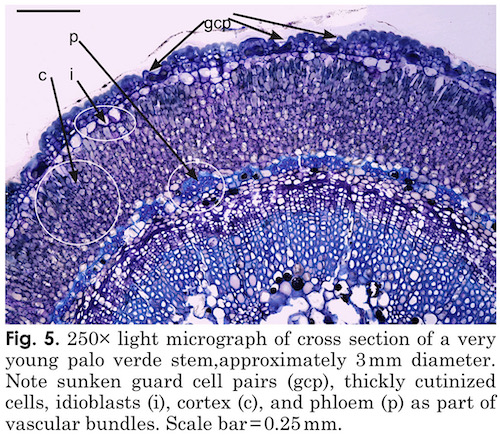
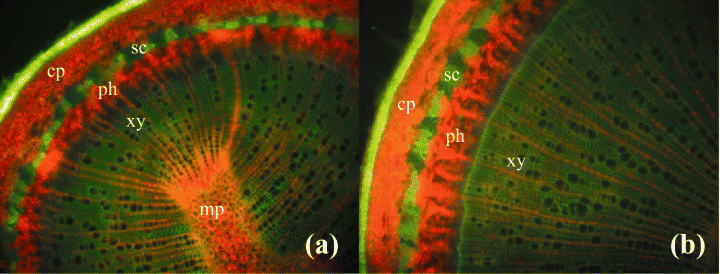
The Palo Verde tree is another very interesting example of C4 photosynthesis in the petioles and trunk and give us some very interesting insight into how light enters an otherwise opaque structure. The epidermis of leaves and stems are heavily cutinized and hair cells are abundant on stems and leaves but rarely survived processing for microscopy. Stem stomata are sunken, (figs. 5, 6) and are arranged randomly along the stem. The sunken and heavily cutinized stoma would prevent unnecessary water loss during hot, dry periods. Sunken guard cells present on palo verde stems are thought to provide an advantage here for two reasons. Most plants feature guard cells on the surfaces of leaves which assist in gas exchange and reduction of water loss. But stem stoma are rare in trees, lenticels take the place of stomates on stems of most woody plants. The fact that guard cells are present and sunken into the stems of palo verde can be considered a fitness feature, because, even on most leaves, guard cells are not sunken in this way. Hair cells (trichomes), present in significant numbers on palo verde leaves, petioles, and stems are thought to interrupt air-flow over plant surfaces, to allow water absorption via contact, and to reduce loss of water in arid and other biomes.
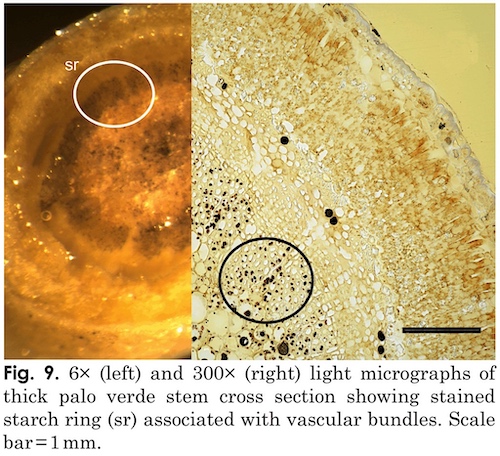
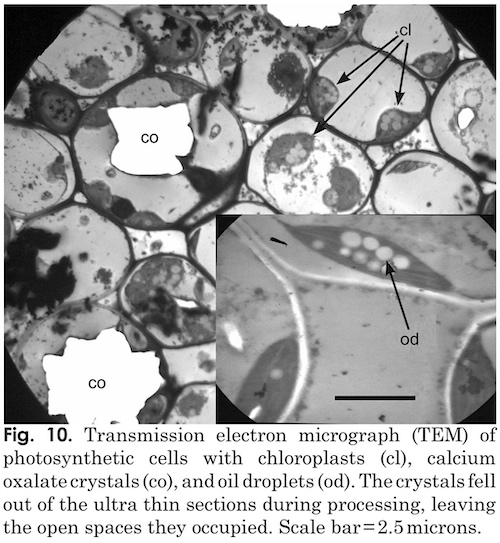
Xylem tissues were thickly lignified. Starch, stained darkly with iodine, was present in abundance and was well distributed across phloem and xylem; however boundaries between these tissues were difficult to identify. Starch grains were highly concentrated in association with a circumferential ring of phloem (fig. 9). Many oil droplets were found deposited within photosynthetic cells and especially in chloroplasts (fig. 10). Large amounts of calcium oxalate crystals were also deposited throughout the stem, and between stoma and the outer border of the cortex. Photosynthetic cells usually contained crystals of calcium oxalate or lipids, or sometimes both, (fig. 10). Many chloroplasts were often bulging with oil droplets (fig. 10). Calcium oxalate is a common component of plants, and often occurs as long thin and clear crystals. There are enough of them to aid light in penetrating the otherwise opaque plant structures.
Boojum Tree (C3 Leaves and C4 Sarcocaulescent Stem)

Fouquieria columnaris, the Boojum tree or cirio is a tree in the ocotillo family, whose other members include the ocotillos. It is nearly endemic to the Baja California Peninsula. Boojum tree is one of the most bizarre-appearing plants in the world. Shaped like snakey naked candles or upside down carrots, they never fail to interest observers. Boojums are native to the Sonoran Desert on the west coast of Baja California. and one small area on the Sonoran mainland near Puerto Libertad. As with its relative the ocotillo, the small leaves fall early, leaving the greenish stems to carry out food-producing photosynthesis. The swollen trunk base is often hollow provides a habitat for honeybees, the wood is somewhat spongy and retains water. The boojum tree is sometimes planted in southern California and Arizona as a landscape curiosity; small plants can be grown indoors. Studies show the photosynthesis in the leaves is C3. The stem is green and it appears that CO2 produced by photorespiration is trapped in the stem and reused for stem photosynthesis, thus utilizing C4 photosynthesis in a sarcocaulescent stem.



Succulent pachycauls or the sarcocaulescent group are unusual plants whose physiology is incompletely understood. Many examples of this group are found in Africa and the spiny forests of Madagascar, as seen in the examples above. Some authors have tried to categorize stems into three categories, cactus, retinoids (with stomata) and sarcocaulescents (without stomata). This is just wrong on so many levels since stomata are one of the least important things about pachycauls. A clear and concise framework of stem photosynthesis and physiology at this time remains a goal of future research.
Ocotillo (C3 Leaves and C4 Sarcocaulescent Stem)

Ocotillo is not a cactus as some might think. It is a member of a small, unrelated family of primarily South American plants, of which it is the northernmost representative. While semi-succulent and a desert plant, Ocotillo is more closely related to tea and blueberries than to cactuses. It is a member of, Ericales, a small, unrelated family of primarily South American plants, of which it is the northernmost representative. It is found across the Chihuahuan and Sonoran deserts of Texas, New Mexico, Arizona, and a small part of California. Even though it is grown in Nevada, it is not really native to the Mohave desert. I am including it due to the important adaptations of stem photosynthesis. The Ocotillo has adapted to its environment by shedding its small leaves during dry spells (drought deciduous). It can also grow new leaves 5 days after getting water. It has a shallow, but wide root system, which it uses to gather rainwater. It produces food because the Ocotillo can perform photosynthesis during dry spells in its woody stem.

For much of the year, the plant appears to be an arrangement of large spiny dead sticks, although closer examination reveals that the stems are partly green. With rainfall, the plant quickly becomes lush with small (2–4 cm), ovate leaves, which may remain for weeks or even months. This cycle of growing and dropping leaves can and does happen multiple times per year. It is not uncommon to see ocotillo leaf out up to 4 or 5 times between spring and fall. During the rest of the year, ocotillo relies on chlorophyll in its stems for its photosynthetic needs. Interestingly enough, this poses a bit of a challenge when it comes to getting enough CO2. Whereas leaves are covered in tiny pores called stomata which help to regulate gas exchange, the stems of an ocotillo are a lot less porous, making it a challenge to get gases in and out. This is where the efficient metabolism of this plant comes in handy.

All plants undergo respiration like you and me. The carbohydrates made during photosynthesis are broken down to fuel the plant and in doing so, CO2 is produced. Amazingly, the ocotillo (as well as many other plants that undergo stem photosynthesis) are able to recycle the CO2 generated by cellular respiration back into photosynthesis within the stem. In this way, the ocotillo is fully capable of photosynthesis even without leaves. The plant is leafless most of the year because of drought, so the capacity for gas exchange by the stems is essential for their photosynthetic function. With secondary growth, sclerified leaf bases which cover the stem become separated, and a transparent cork forms in the furrows between them. Light is transmitted through the leaf base on the young primary shoot and the furrow cork, but not through the older leaf base. Despite competent chloroplasts, no exogenous CO2 uptake occurs because the cork is impermeable to CO2, and presumably water. The functional significance of competent chloroplasts in stems that do not transfer gas may be the production of high energy compounds for metabolism, the recycling of internally generated respiratory CO2, or it may simply be a relict feature in this species of the Fouquieriaceae.

Under stressful conditions, stems increase their proportional contribution to canopy carbon gain compared with leaves. The Fabaceae and Asteraceae may be the families with the greatest diversity of stem photosynthetic species. Corticular photosynthesis occurs in desert and temperate forest species, probably with the primary purpose of recapturing respired CO2 from other organs. CAM stem photosynthesis occurs in succulent species to maximize water use efficiency. Stem photosynthesis is similar to C3 leaf photosynthesis and occurs in species inhabiting a diversity of high-light sites.

Although net photosynthetic uptake of CO2 is rarely found, stem internal re-fixation of CO2 in young twigs and branches may compensate for 60–90% of the potential respiratory carbon loss. Isolated chlorenchymal tissues reveal rather high rates of net photosynthesis (being up to 75% of the respective rates for leaf photosynthesis). Corticular photosynthesis is thus thought to be an effective mechanism for recapturing respiratory carbon dioxide before it diffuses out of the stem.

One of the most striking aspects of ocotillo life is how quickly these plants respond to water. As spring brings rain to the desert, ocotillo respond with wonderful sprays of bright red flowers situated atop their spindly stems. These blooms are usually timed so as to take advantage of migrating hummingbirds and emerging bees that pollinate their flowers. The collective display of a landscape full of blooming ocotillo is jaw-droppingly gorgeous and a sight one doesn’t soon forget. It is as if the whole landscape has suddenly caught on fire. Indeed, the word “ocotillo” is Spanish for “little torch.”
Nevada Ephedra (C3 Retamoid Stem)

Ephedra is a genus of gymnosperm shrubs. Nevada ephedra was used for food and medicine by indigenous people and for tea by Mormons. Its range extends west to California and Oregon, east to Texas, and south to Baja California, including areas of the Great Basin, Colorado plateau and desert Southwest. It is found in rocky and sandy soils, generally in areas without trees. The stems are green with stomata and utilize C3 carbon fixation. The tiny leaves (it really does not have leaves) are opposite or whorled. The scalelike leaves fuse into a sheath at the base and this often sheds soon after development.
Desert Milkweed (C3 Retamoid Stem)



Asclepias subulata is a species of milkweed known commonly as the rush milkweed, desert milkweed or ajamete. This is an erect perennial herb which loses its leaves early in the season and stands as a cluster of naked stalks. Atop the stems are inflorescences of distinctive flowers. Each cream-white flower has a reflexed corolla that reveals the inner parts, a network of five shiny columns, each topped with a tiny hook. The fruit is a pouchlike follicle that contains many flat, oval seeds with long, silky hairlike plumes. This milkweed is native to the desert southwest of the United States and northern Mexico. I assume this is a C3 photosynthesis plant although I can find no specific study confirming this. I have personally seen this plant in washes and in riparian transition zones although it is reported on dry hillsides and plains.
Brown-Plume Wire-Lettuce (C3 Retamoid Stem)


Stephanomeria pauciflora is a species of flowering plant in the aster family known by the common names brownplume wirelettuce, few-flowered wirelettuce, and prairie skeletonplant. It is native to the southwestern United States and northern Mexico, where it grows in many types of habitat, including many desert areas, woodlands, and plains. It is a perennial herb or bushy subshrub producing one or more sturdy, stiff stems with many spreading branches, taking a rounded but vertical form. The leaves are mostly basal and ephemeral, with smaller, scale-like leaves occurring on the upper stem. Flower heads occur at intervals along the mostly naked stems, especially near the tips. Each has a cylindrical base covered in hairless phyllaries. It contains 3 to 6 florets, each with an elongated tube and a flat pink ligule. The fruit is an achene tipped with a spreading cluster of plumelike pappus bristles. These are usually brownish, but are sometimes white. The specific epithet pauciflora, refers to the Latin term for ‘few flowered’. This is a C3 plant for both leaves and stem based on carbon isotope ratios of 22.3 and 23.3 respectively (Ehleringer et al. (1987)).
Desert Trumpet (C3 Leaves and C3 Retamoid Stem)



Eriogonum inflatum, the desert trumpet, is a perennial plant of the family Polygonaceae. The plant possesses very small yellow or pink flowers and an inflated stem just below branching segments. It is found in the Mojave Desert and other deserts. The swelling of the stems was assumed to have been influenced by the presence of gall insects, most notably of the genus Odynerus. The female wasp produces a small hole on the inflated portion of the desert trumpet, packs the cavity with larvae and lays her eggs upon them, providing a food source, and a protected environment for the offspring. Irritation caused by this process was said to have enlarged the cavity over time. Recently, research performed by a world authority on the genus Eriogonum, Dr. J. L. Reveal of the University of Maryland, College Park, revealed the swollen stem of Eriogonum inflatum is due to high concentrations of carbon dioxide in the solid stem and seems to be related to gas regulation and stem photosynthesis. Inflated stems were found to contain extraordinarily high internal CO2 concentrations (to 14000 μbar), but fixation of this internal CO2 was 6–10 times slower than fixation of atmospheric CO2 by these stems. Although the pool of CO2 is a trivial source of CO2 for stem photosynthesis, it may result in higher water-use efficiency of stem tissues. Green stems accounted for 66–77% of whole plant photosynthetic surface area when integrated over a developmental cycle, whereas only 40–67% of the yearly transpirational water loss could be attributed to stems. Stems were found to have lower nitrogen and chlorophyll contents than leaves, and lower stomatal conductance under all physiological conditions encountered. Eriogonum is found throughout Arizona and entire southwest with 58 species in Arizona, 115 species in California, 36 species in New Mexico, 79 species in Nevada and 67 species in California. I assume this is a C3 photosynthesis plant although I can find no specific study confirming this.
Skeleton Weed (C3 and C3 Retamoid Stem)

Eriogonum deflexum is a species of wild buckwheat known by the common names flatcrown buckwheat, flat-top buckwheat, and skeletonweed. This plant is native to the southwestern United States and northwestern Mexico, where it is common and grows in a variety of habitats, especially desert scrub. It is somewhat weedy where it is most abundant. This is an annual which varies in size from small patches on the ground to tangled bushes approaching two meters in height; it may be dense or thin and spindly. This is a brown or greenish weedy-looking herb with a many-branched stem. The leaves are located at the base of the plant and are rounded and woolly and one to four centimeters long. Small clusters of flowers appear at intervals along the branches with each flower only one to three millimeters wide and white or pinkish in color. I took this photo in late December and as you can see it is in flower. The structure of this plant demonstrates two very different xerophytic adaptations. First the broad leaves are set very close to the soil. Here, next to the ground, there is a boundary layer of slower moving, slightly more humid air. In this microenvironment the leaves can better perform their photosynthetic work while minimizing water loss. Surprisingly, there is competition for pollinators at critical times and as a result tall flowering stems that can push above the competition have been selected for. Making double-use of this scaffolding for photosynthesis, skeleton weed inflorescence stems are smooth, green and leaf-free. Thus above the soil boundary layer, where drying winds are persistent, skeleton weed minimizes water loss by minimizing leaf surface area while keeping overall surface area high to improve heat dissipation and thus increase photosynthetic efficiency. I assume this is a C3 photosynthesis plant although I can find no specific study confirming this.
Parasitic Plants
Most plants also have other pigments: carotenoids, which usually appear yellow to orange, and anthocyanins, which are red to purple. One pigment usually dominates. So a plant with red leaves probably has higher than usual amounts of anthocyanins. But chlorophyll is still present and at work. In photosynthesis, carotenoids help capture light, but they also have an important role in getting rid of excess light energy. When a leaf is exposed to full sun, it receives a huge amount of energy; if that energy is not handled properly, it can damage the photosynthetic machinery. Carotenoids in chloroplasts help absorb the excess energy and dissipate it as heat. I included this material on carotenoids since Mohave desert plants are sometimes red or brown.
Desert Mistletoe (Hemi-Parasitic)


Phoradendron californicum, the desert mistletoe or mesquite mistletoe, is a hemiparasitic plant native to southern California, Nevada, Arizona, Sonora, Sinaloa and Baja California. It can be found in the Mojave and Sonoran Deserts at elevations of up to 1400 m (4600 feet). The leaves are reduced to scalelike bracts and are mostly connate (pairs are joined). In the trees and bushes of this desert region, desert mistletoe often looks like a dense nest or hive. In reality, desert mistletoe is a dense cluster of brittle, jointed green stems that are primarily leafless. The clusters tend to hang down from the host and can grow to lengths of 36 inches (1 m) in six to eight years. Desert mistletoe takes water and minerals from its host plants but it does its own photosynthesis, making it a hemi-parasite (partially parasitic).

The seeds of the fruit are covered with a gelatinous material that makes them extremely sticky. They are a favorite food for the birds of the desert that often get the sticky seeds stuck to their beaks or feet. As they wipe their beaks and feet clean on another branch, or even another tree, the mistletoe seeds now become planted on a new potential host. Since the seeds also have a tough seed coat, they easily pass through the bird’s digestive system and become deposited onto new branches through their droppings. Desert mistletoe plants, but not the berries, contain phoratoxins which can easily lead to death via slowed heart rate, increased blood pressure, convulsions, or cardiac arrest.


The Phainopepla (Phainopepla nitens), commonly known as a Silky Flycatcher, is a small, desert bird that seems to have a special relationship with desert mistletoe. The black, male Phainopepla looks like a miniature cardinal. The mistletoe berries are the bird’s main source of food in the winter. As such, the Phainopepla is one of the birds primarily responsible for the distribution of desert mistletoe seeds.
Dodder (Parasitic)

Cuscuta californica is a species of dodder known by the common names chaparral dodder and California dodder. It is native to western North America. Cuscuta californica is a parasitic vine which climbs other plants and takes nutrition directly from them via a haustorium. The dodder resembles a pile of yellow-orange straw wrapped tightly around its host plant. It is mostly stem; the leaves are reduced to scales on the stem’s surface, since they are not needed for photosynthesis while the dodder is obtaining nutrients from its host. It bears tiny white flowers which are only about 3 millimeters wide, and fruits which are even smaller.
As always I hope you enjoyed the post, if you have read to this point you deserve applause.
References:
Stem-Succulent Trees from the Old and New World Tropics
SCHAEDLE, M. 1975. Tree photosynthesis. Annu. Rev. Plant Physiol. 26: 101–115.
COMPARATIVE GAS EXCHANGE OF LEAVES AND BARK IN THREE STEM SUCCULENTS OF BAJA CALIFORNIA
Tobacco C3 Leaves and C4 Stems. Julian M. Hibberd & W. Paul Quick
The photosynthetic response of tobacco plants
Stem photosynthesis in a desert ephemeral, Eriogonum inflatum
Stem photosynthesis and hydraulics are coordinated in desert plant species
A Light and Electron Microscopic Study of the Palo Verde Tree of the Desert Southwest
Calcium oxalate crystal formation and function in plants
Contribution of stem CO2 fixation to whole-plant carbon balance in
nonsucculent species
The Advantages of Having Green Stems in Arid Environments
A Virtual Tour of Major Weed Plant Families
Anatomy of Photosynthetic Old Stems of Nonsucculent Dicotyledons from North American Deserts
Unified representation of the C3, C4, and CAM photosynthetic pathways T with the Photo3 model